【アメーバブログだけの追記】
この投稿は、記事筆者が運営するウェブサイト中の内容が、記事筆者の死後には、運営ウェブサイト自体が確実に閉鎖されるため、アメーバブログに記事筆者死後用に保存したものです。また、再投稿ではなく、前回のアメーバブログ投稿内容にかなりの加筆修正を行っています。
このアメーバブログには、日韓カップルというジャンルすら存在しますが、私としては、日本人のみならず全人類が知るべき科学的事実であると確信しています。
アメーバブログの日韓ジャンルに投稿されている方々が、万一読まれた場合には、さぞかし気分が悪いことでしょうが、全て根拠論文や根拠となるデータを明示している点をご理解ください
(ページ最下部にアメーバブログだけの追記をしました。)
韓国人DNAの特異性と朝鮮半島独特の奴隷制による意図せざる近親相姦
複数の集団遺伝学論文により、韓国人(=朝鮮人)DNAの特異性は疑問を差しはさむ余地がほぼありません。 高麗末期から李氏朝鮮時代を通じて700年も維持され続けた朝鮮半島独特の奴婢制=奴隷制=「人口の30%にも及ぶ同じ民族即ち同胞を奴隷として売買・相続の対象とするという 人類史上朝鮮半島にしか存在しなかった極めて特異な制度」こそが、膨大な数の「意図せざる近親相姦」の集団的・累積的・継続的発生により、朝鮮人遺伝子プールに重大な影響を及ぼしたものと推測されます。
同父異母者間の交配は、通常の人類社会では非常に稀なケースなのですが、日韓併合前の朝鮮半島では「同胞奴婢制=同胞奴隷制」の故に、社会制度上、同父異母者間の超近親交配(=近親相姦)が不可避でした。なお、日韓併合(=日帝強占→韓国の表現)により朝鮮半島独特の奴隷制は完全に根絶されました。

集団遺伝学論文データから見て朝鮮人DNAの特異性は明白です
複数の集団遺伝学論文から朝鮮人=韓国人D NAの遺伝的構造の特殊性は疑問の余地が全くありません。

上の図は下記の集団遺伝学論文の附属資料から。
①Whole genome sequencing of 35 individuals provides insights into the genetic architecture of Korean population
Wenqian Zhang et.al
21 October 2014
(注)1
上記論文が指摘する朝鮮人の遺伝的特質・医療上の注意喚起は、次のとおり
*朝鮮人固有の変異は、極度に集団内の一部の者に集中している。(サンプル数35名の8%程度の者は極度に朝鮮人固有の変異を大量に有している)

*集団として朝鮮人固有の変異は、集団内から消滅するはずの非同義変異が他民族集団に比べ高い比率である
*従って、個別化医療に際しては、朝鮮人に対しては特段の配慮を払うことが必要である
*論文執筆者に韓国人又は韓国系アメリカ人は全くおらず、事実上はアメリカ食品医薬品局(FDA)によって書かれた論文である、
*この論文の内容はあまりにも重大なため、
韓国人DNAの特異性を明確に示す論文(アメリカ食品医薬品局)の内容紹介として、この論文だけを対象に解説記事を書いています。
②Korean National Standard Reference Variome database of whole genomes with comprehensive SNV, indel, CNV, and SV analyses
Jungeun Kim et. al
Nature scientific reports 04 April 2018
韓国人による論文②では、朝鮮人固有の変異が、集団内の一部の者に極度に集中している点に関して同様の結論 であることに加え、集団内の変異分布が他の民族集団と大幅に異なることを本文中で明記している。これら2点は論文①の内容と完全に合致する

【上記論文から引用】
Surprisingly, however, roughly half of the variants in ‘1000GP low frequency’ were classified as ‘frequent in KoVariome’. This indicates that there exist a significant population specific biases for common and uncommon variants.
【英文の趣旨】 しかしながら、驚くべきことには、(国際的なDNA分析プロジェクトである)1000ゲノムプロジェクトで低い頻度に区分されたバリアンツ(変異)のおよそ半分が、 KoVariome(と名付けた韓国のDNA分析プロジェクトでは)高い頻度であった・このことは、ありふれたバリアンツ及びレアなバリアンツ(の分布に関して、韓国人には)重大な集団固有の偏り(=バイアス)が、存在することを示している
(注) 2
③Extensive genomic and transcriptional diversity identified through massively parallel DNA and RNA sequencing of eighteen Korean individuals
YS Ju et.al
Nature Genetics 03 July 2011
韓国人3名以上のサンプル数で初めて出された上記全ゲノムシーケンス論文では、韓国人の非同義一塩基変異が他の民族集団 に比べ、著しく高い頻度を示している旨を本文中で明言しており、論文①と完全に合致する。
【上記論文から引用】
A subset of the nsSNPs showed remarkably high allele frequencies among the Koreans studied compared to other populations, including Europeans and west Africans represented in the HapMap project.
【英文趣旨。カッコ内は英文の意味が分かるように筆者補足】
(分析対象の)部分集合としての韓国人の非同義一塩基多型(=nsSNPs)の頻度は、ハップマッププロジェクトにおけるヨーロッパ人及び西アフリカ人をはじめとする民族集団との比較において、顕著に高い対立遺伝子頻度を示していた。
しかしながら、上記全ゲノムシーケンス諸論文のサンプル数は、①の論文35名、②の論文50名、③の論文10名 と少ないサンプル数に過ぎなかった
このため、筆者が1000名以上のサンプル数の集団遺伝学論文を探した結果として、大規模サンプルでも、非同義変異比率が他の民族集団に比べて高いという朝鮮人D NAの特性は 完全に確認出来てます。
④A database of 5,305 healthy Korean individuals reveals genetic and clinicalimplications for an East Asian population
Jeongeun Lee et al.
Nature 02 November 2022
韓国人対象5,000名を超える大規模サンプル数の上記韓国人論文では、本文中で非同義変異が他民族集団に比べ高い比率 である旨を一切言及していない。しかし、下図が掲載されている

論文⑤A global reference for human genetic variation The 1000 Genomes Project Consortium
The 1000 Genomes Project Consortium
Nature 2015 Sep 30
1000ゲノムプロジェクトフェーズ3の分析対象集団(韓国人は含まれない)

論文④と⑤のエクソン領域のゲノム解析深度(coverage depth)に大きな差はないことから(論文④は最低でも40x、論文⑤は平均67x)、
対比として、singleton(個体固有)のみを対象に、上記1000ゲノムプロジェクト結果発表論文⑤と論文④を突合することは可能であり、
朝鮮人は非同義変異が他民族集団に比べ、朝鮮人固有の変異のみではなく、エクソン領域全体で非同義変異が高い比率であることが大規模サンプルにて完全に確認しえ、論文①及び③と完全に合致する。
論文⑤の表1からsingleton(個体固有)のみを抽出したデータと論文④付属資料データから作成した表が下記
二つのデータを統合・比較すると、他の民族集団比べて、朝鮮人の場合には非同義変異比率が17%程度も高い。 コード領域(=エクソン)に生じる非同義変異は、過去1000年に限れば、ほぼ全てが負の自然淘汰の対象である遺伝子変異であり、集団内からは「消え去るべき」遺伝子変異であることは 絶対に間違いありません。従って、他の集団に比べて非同義変異比率が高いことは、朝鮮人遺伝子の恐ろしいほどに重要な集団特性です。
朝鮮人=韓国人が突出してエクソン領域の非同義変異比率が高いことは、過去1000年以内に朝鮮半島において何らかの特殊な事象が生じたことを意味する。
言い換えれば、20世紀初頭以降朝鮮半島の人々のDNAは正常化の方向にあるものの、非同義変異比率の低下はゆっくりとしか進まないため、 完全な正常化には、長い世代(500年?)を経る必要がある。
| singleton= 個体固有 |
非同義 変異 |
同義 変異 |
比率 |
| アフリカ大陸 661名 論文⑤データ |
139,000 | 78,000 | 1.78 |
| 南北アメリカ大陸 347名 論文⑤データ |
121,000 | 67,000 | 1.80 |
| 東アジア 504名 論文⑤データ |
144,000 | 79,000 | 1.82 |
| ヨーロッパ 503名 論文⑤データ |
116,000 | 59,000 | 1.96 |
| 南アジア 489名 論文⑤データ |
144,000 | 78,000 | 1.84 |
| 韓国人 5305名 論文④付属資料 |
274,693 | 127,844 | 2.15 |

次に、何故、韓国人D NA=朝鮮人D NAにのみこの様な奇妙な集団的特性が生じたのか?
初期原因としての高麗前期の近親婚・近親相姦の蔓延
高麗時代(918年~1392年)初期における朝鮮半島では、下記論文に従えば、近親相姦(父が同じで母が異なる関係にある兄弟姉妹の間での結婚)と近親婚が多かったことは間違いない。 にもかかわらず、この重大な歴史的事実は、日本では全く知られてはいない。
論文 1
세기 高麗 王室의 構造와 編制 :
The Structure and Organization of the Goryeo Royal Family in the 10th-13th century
황향주 2022
【ハングルのグーグル機械翻訳による】
太祖は生前異腹兄妹関係の王子と王女を婚姻させ、王室の禁婚範囲を同母兄弟姉妹に最小化した。 以後、高麗朝王室では遠親から近親に行くほど婚姻を好む傾向が現れ、王室女性に王室男性との婚姻を強制する內婚規範が成立した。 太祖の子ども世代以降、王室では両親の血統を選び、メンバーを階序化した。親の內婚の可否によって、太祖の孫娘たちには王位継承権に関与する資格が差別的に与えられた。
にわかには信じがたいことであるが、この論文1の記述が正しいのであれば、初代高麗王である王建は、近親相姦を命じている。論文1によれば、その目的は王妃の外戚が跋扈することを防止することにあったという。
高麗王家では、モンゴル軍の朝鮮半島征服以前(=忠烈王以降はモンゴル帝室から王妃を迎えるのが慣例)は、完全な族内婚が実施されていたことは間違いない。
李氏朝鮮時代に編纂された高麗史によっても、高麗前期の近親婚の蔓延は確認できます。
高麗王家及び高麗支配階層の近親相姦・近親婚の慣習
高麗史(紀伝体)及び高麗史節要(編年体)における近親相姦などを記述した箇所の漢文原文は下記の通り。
*志卷第十五 日付1058年06月未来
從之 穆宗・宣宗、同父異母也。
漢文趣旨
第7代高麗王(在位:997年 - 1009年)穆宗と第13代国王(1049年10月9日 - 1094年6月17日)は、同父異母である=父が同じで母が異なる状態で生まれた子である(=近親相姦の結果、生まれた子である)
*志卷第二十九 日付1085年04月未賞
宣宗二年四月判、「同父異母姊妹犯嫁所産、仕路禁錮。」
漢文趣旨
同父異母の子=近親相姦の子は、任官することを禁ずる
【この記述は、高麗王家にとどまらず、高麗支配階層に近親相姦が極めて多かったことを明確に示します。なお、高麗史は中国正史を真似て作られた高麗時代の朝鮮半島の正史です。】
では、近親相姦を含めた近親婚の集団遺伝学から見た影響は?詳細は注3
集団遺伝学では、inbreeding load と呼ばれています。意訳すれば、(ヒトの場合には)近親相姦・近親交配による遺伝的負荷という趣旨であり、分かりやすく言えば、近親婚を繰り返しているヒトの集団では、「生殖可能年齢まで正常に成長できる子孫の数」が減少してしまう、知能が低下するという経験的によく知られた結果が生じます。詳細は、(注)3
inbreeding load(=遺伝子面)は事実上、inbreeding depression(=形質面)と同義であり、ヒトを含めて全ての高等生物に当てはまる現象です。
一般的には、近親交配によるホモ接合対立遺伝子比率の増加を原因とする子孫の環境適応能力の低下です。このため、高等生物では本能的に近親相姦・近親婚を避けようとします。例えば、乱婚制のチンパンジーでは雌が群れを出て他の群れに移る父系社会であり、二ホンザルの場合はオスの子供は4~6歳頃生まれた群れを出てよその群れに入る母系社会です。
複数の集団遺伝学論文が明確に示す現在の朝鮮人=韓国人D NAの特異性が生じた原因の一つとして、高麗時代前期の高麗支配階層における近親相姦、近親婚の蔓延による遺伝的重荷の影響の残滓があると推測される。
決定要因としての李氏朝鮮における奴婢制=奴隷制
李氏朝鮮について、フランス人のC・ダレは次のように明言している
「朝鮮事情」 C・ダレ 東洋文庫
P267~268
朝鮮人は、男女とも、生まれつき非常に熱情的である。しかし、真の愛情はこの国には全く存在しない。彼らの熱情は純粋に肉体的なものであって、そこのは何ら真心がない。 彼らは自分自身を満足させるため、手の届く対象にはなんにでも飛びつくあの動物的な欲望、獣的本能以外は知らない。従って、風紀の腐敗は想像を絶し、「人々の過半数は、自分たちの真の両親を知らない」と大胆に断言さえできる
しかし、ダレは朝鮮半島独特の奴婢制=奴隷制の遺伝的観点からの危険性に気づいてはいない。
かつての南北アメリカ大陸に広範に存在した黒人奴隷制とは異なり、朝鮮半島では完全に同じ民族を奴隷として売買し、両班の相続対象としていた。
このことは、通常の近親相姦・近親婚と全く異なり、
「意図せざる近親相姦・近親婚」の危険性を極度に高める
例えば、両班がその所有する奴婢女性に女児を産ませ後、同じ両班男性の妻が男児を生み、その後成長した両班の男児が(両班は奴婢女性と自由に性交できるが故に)同父異母の成長した奴婢女児に子供を産ませるケースが非常に多数存在しうる。
両者は同父異母の兄弟姉妹であり、この両者の間に子が生まれた場合、完全な意図せざる近親相姦である。
同胞を奴隷としていた朝鮮史の致命的な欠点である。
人類史において朝鮮以外では、奴隷は全て購入奴隷・戦争捕虜奴隷であり、奴隷制内部で奴隷が継続的に再生産されることはなかった。しかし、朝鮮独特の同胞奴隷制は奴隷を再生産しうるという利点を有している反面、極めて多数の超近親交配(=同父異母の子どうしの交配の発生)を朝鮮人集団内に 集団的・累積的に発生させざるを得ない
パターン①奴隷男女間
Changes in informal society and slavery during the Chosun-Era in Korea
Heeho Kim
Nature humanities and social sciences communications
19 October 2022
上記論文によれば、韓国慶尚道のある両班家に残る記録から、1801年時点で、女性奴隷(=婢)を41名、男性奴隷(=奴)を32名所有していたことが確認でき、 郷班・土班と呼ばれる自称両班を除く正規の両班家は、平均して10人~50人前後の奴婢を所有していたのでは?と思われます。

奴隷主である両班にとって、一番頭の痛い問題は、所有する奴隷=奴婢間の近親交配累積による所有奴隷数の減少であったと思われます。 古代より家畜を所有する遊牧民は、この問題を良く知っていたことは間違いなく、奴隷主の両班も確実に知っていたと推定されます。
この問題解決方法は明らかではないのですが、下記のオランダ人の記述からして、両班同士で子供を交換又は売却後に購入していたと思われます。 即ち、パターン①は数としては少なかったのでは?と思料されます。
朝鮮幽囚記 ヘンドリック・ハメル(17世紀の朝鮮に関する記述)
p50 奴隷やそれに類する人々は、ほとんど子供をかまいつけません。それは子供たちが仕事ができるようになると、彼らの主人がすぐに人をやって、彼らを引き取らせるからです。
パターン②奴隷主の両班男性と奴隷女性間

集団遺伝学論文データに基づく同胞奴隷制による近親交配の形跡
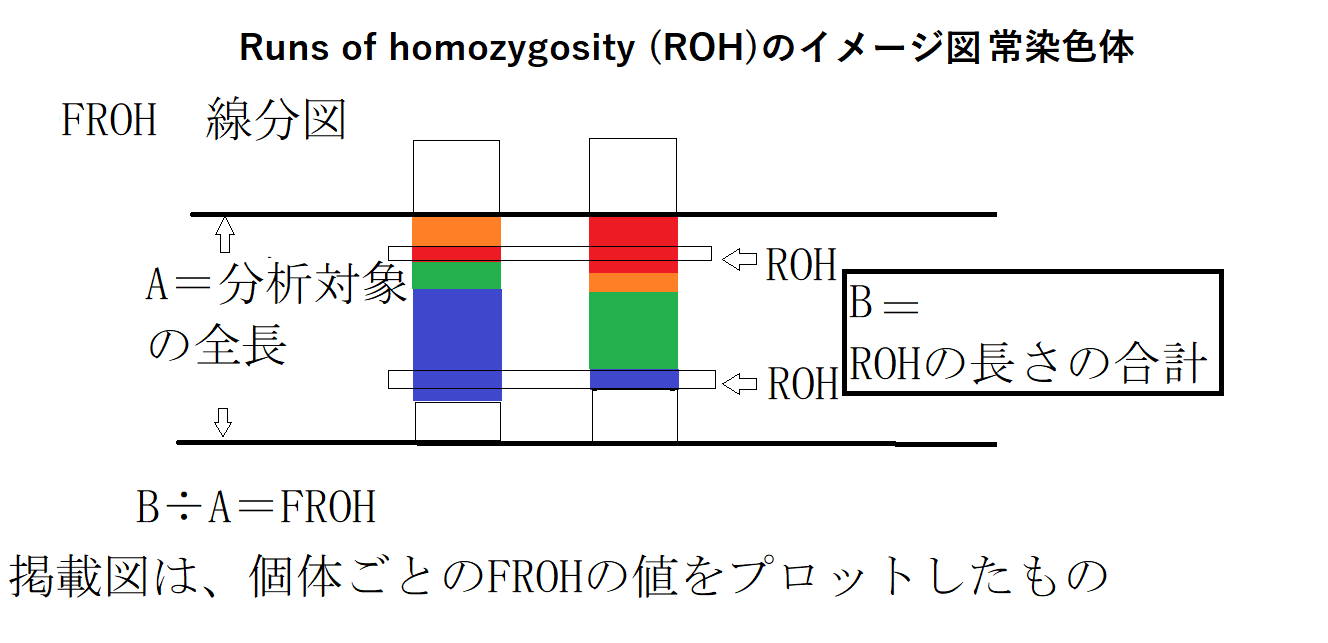
ROH(=run of homozygosity=ホモ接合性連続領域)とは、ゲノム上の染色体対(両親から受け継いだ対の染色体2本)において、 同一の配列(ホモ接合体)が連続して存在する領域のことです。近親交配やボトルネックにより、共通の祖先から受け継いだ相同な領域が連続することで生じます。
10Mb以上の長さのROH(ホモ接合体領域の合計値)が検出された場合には、近親交配とみなす場合が多く、いとこ婚(1C=first cousin)、二重いとこ婚(2C=second cousin=集団遺伝学では近親交配として扱う)などの場合に生じます。
親子間、全血兄弟姉妹間(full sibling)、半血兄弟姉妹間(half siblings=同父異母)のような超近親交配(=近親相姦)の場合には、常染色体の非常に多数の領域でROH=ホモ接合性連続領域が生じます。 過去において一度でも生じた超近親交配により発生した数多くのROH=ホモ接合性連続領域は、次世代にも切り刻まれた状態で受け継がれていきます。
即ち、ROH線分図から、分析対象集団の現在及び過去の状況を推定することが出来ます。
(例)いとこ婚が多いパキスタン人・インド人の場合には、長いROH線分図を示す
ROH線分図は集団遺伝学では、最も有効かつ正確な現在及び過去の近親交配・founder effect・ボトルネック効果等々を示す分析手法で、ROH線分図には3タイプあります。 詳細は、注(4)ご参照。
なお、現時点に限定(一世代前)での近親婚検出には別の手法もあります、下記論文では韓国人をサンプルには含みませんが、全世界の2497名をサンプルとして、 8名が親子間の子、4名が兄弟姉妹間の子であることを検出しています。しかし、高麗末期から李氏朝鮮時代における同胞奴隷制により、朝鮮半島では 半血兄弟姉妹間の子(同父異母間の子)が、人類の歴史上、集団として唯一と言えるほど異様に高い比率で生まれていたことは確実です
High level of inbreeding in final phase of 1000 Genomes Project
Steven Gazal et al.
02 December 2015
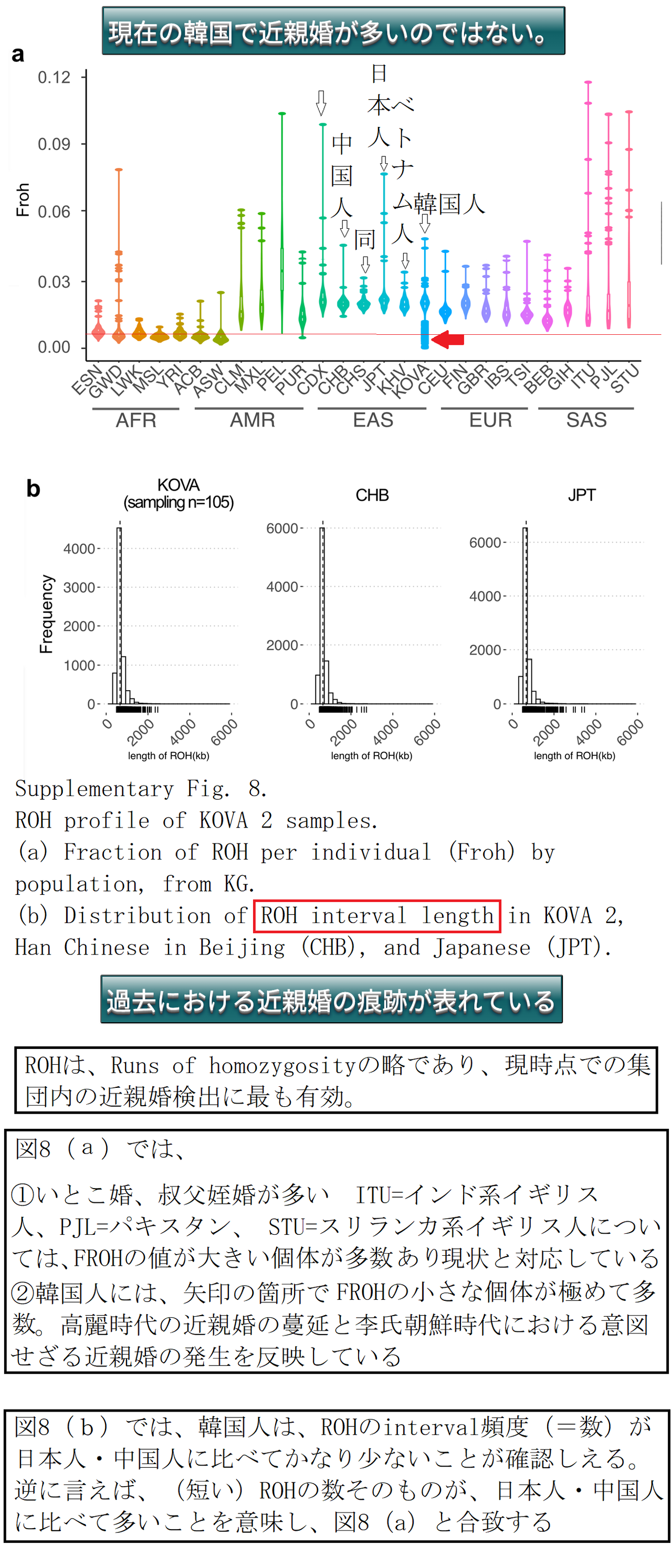
その痕跡は、下のROH線分図に明確に現れています。
ROH線分図中で線分の長い集団は、GWD=アフリカ、ガンビアのマンディガ族、PELはペルー人、 CDXは中国の少数民族のダイ族、ITUは、
イギリスに住むインド人中のテルグ語を話すインドし、PJLはパキスタン人、STUはスリランカに住むタミル人です。
GWD(ガンビア)以外のアフリカの諸民族集団が短い まとまった状態であるのは、他の地域の集団よりも遥かに長い期間(少なくとも10万年以上)現生人類として生存してきたこと(=人類のアフリカ単一起源説)、及び、直近時点でもこれらのアフリカ諸集団内に、いとこ婚等の近親交配がないことを示しています。
これに対して、韓国人は、明らかに非常に奇妙な状態です。日本人もROH線分が長いですが、これは日本では、いとこ婚への忌避慣習はあっても、二重いとこ婚(2C=second cousin者間の交配は集団遺伝学では近親婚と扱われます)への忌避慣習は がほとんど見られないことによるものではなかろうか?と思われます。
現在の韓国で近親婚は民法上厳重に禁止されており、建前上は、8親等以内の者の婚姻は禁止されています。 しかし、現実には8親等以内ではないことの戸籍上の証明は、婚姻届けで提出の際には求められていないようです。現在の韓国で近親婚が多いことを示す図ではありません、勘違いしないでください
つまり、下のROH線分図中の韓国人だけがハッキリと示す奇妙な状態は、直近時点ではなく、もっと前の何らかの極めて特殊な要素が原因であることは絶対に確実です

では、このような朝鮮人即ち韓国人D NAの特異性が明確に発現しているデータが存在するのか?
2024年12月18日フランス系通信社であるAFPは次の驚愕する記事を配信しました。以下は、記事全文です。(注)5
韓国の成人「言語・数理・問題解決」能力、残念ながらOECD平均を下回る
2024年12月18日 5:00 発信地:韓国 [ 韓国・北朝鮮 ]
【12月18日 KOREA WAVE】韓国成人の言語能力、数理能力、問題解決能力が経済協力開発機構(OECD)の加盟国平均を下回るという調査結果がOECDの「国際成人能力調査(PIAAC)」で明らかになった。
調査は、16~65歳の成人を対象に言語能力、数理能力、適応的問題解決力などの情報処理スキルを測定し、日常生活や職場での能力活用レベルを比較するもの。10年ごとに実施されている。 今回の第2サイクル調査では、前回の23カ国から8カ国増えた31カ国で約16万人が参加し、韓国からは6198人が調査に応じた。
それによると、韓国成人の平均点数は言語能力249点、数理能力253点、適応的問題解決力238点であり、いずれもOECD平均(言語能力260点、数理能力263点、適応的問題解決力251点)を下回った。
前回の調査結果と比較可能な領域でみると、韓国の言語能力は273点から249点へ、数理能力は263点から253点へ低下した。
韓国教育省は「第1サイクルと第2サイクルを比較した場合、言語能力の平均点は24点低下したが、調査対象27カ国のうち、平均点が上昇したのはフィンランドとデンマークのみだった」と説明。一方、数理能力の低下(10点)は統計的に有意な変化ではないと付け加えた。
調査対象を16~24歳に限定した場合、韓国の言語能力は276点、数理能力は273点で、それぞれOECD平均(言語能力273点、数理能力272点)とほぼ同等の結果となった。
学歴不一致に関する調査では、韓国の16~65歳成人のうち、実際の学歴が仕事で求められる学歴と一致する割合は65.0%だった。一方、学歴過剰は31.3%、学歴不足は3.7%で、OECD平均(学歴過剰23.4%、学歴不足9.5%)より学歴過剰の割合が高い。
スキル不一致では、実際のスキルが業務で求められるスキルと一致する割合が65.1%。スキル過剰は23.9%、スキル不足は11.0%で、OECD平均(スキル過剰26.1%、スキル不足9.6%)と比較するとスキル不足の割合がやや高かった。
記事中のOECDによる加盟各国の成人対象の各分野のスキル調査(=事実上の知能調査)は、10年に一度しか実施されません。各国とも対象サンプル数は非常に多く、 例えば韓国の場合にはサンプル数は6000名を超えており、サンプル誤差がほとんど生じえないデータです。反面、ウェブ上では詳細内容は公表されておらず マスコミ関係者が報道する場合ですら、OECD事務局に開示請求して初めてデータを入手しえます。下記画像はAFP及び朝日新聞記事から記事筆者が作成しました

記事筆者はPIAACデータそのものを入手しえないため、Gemini(=グーグルの対話型AI)を通じて、次のデータを入手しました。
若年層(25-34歳付近)のスキルスコア比較(2024年発表データ)

Geminiに対して、趣旨として
「日本と韓国の大学進学率を比べると韓国の方がはるかに高い、即ち、少なくとも両国の教育環境はほぼ同一、又は、韓国が上回っているはずだ! にもかかわらず、何故、これほどの極端な差が出るのか?ありえない!遺伝的要因である!」
と問うた結果、Geminiは上記データとともに下記の通り返答抜粋してきました。
次に大学卒業者だけで比較して欲しいとの要求にGeminiは次のデータを返答し、枠内の返答抜粋をしてきました。

実は、このタイプの問題に関して、教育等の環境ではなく、遺伝的要素が原因であると主張することは民族差別に直結するため、(特に研究者の場合には)ダブー中のタブーであり、 絶対にそのようなことは口が裂けても言いません。しかし、記事筆者は画像のとおりハッキリと主張します。恐らくは、この記事も検索には絶対に反映されません。 何故なら、グーグル検索はこのタイプの問題を完全排除する基本設定であるからです。
この投稿が、「韓国人は近親相姦(試腹)で遺伝的に異常」などという空虚な、根拠のない、データがない、非常に分かりやすい、俗説・虚説と異なる点だけはご理解ください。
事実やデータを「知る権利」そのものが失われてしまうと考えます。

朝鮮人DNA(=韓国人DNA)の特異性の発生原因は?(まとめ)
高麗前期における支配階層での近親婚・近親相姦の蔓延
↓
その後のモンゴル軍侵攻による人口の大幅な減少(推計で85%減)
↓
李氏朝鮮時代の「意図せざる近親婚・近親相姦」の発生(この記事)
(注) 1 朝鮮人固有の遺伝子変異が生じている遺伝子リスト
この論文では、韓国人だけが有する韓国人固有の遺伝子変異が生じている遺伝子リストが、付属資料にてエクセル形式で additional_file_7 として公表されている①additional_file_7 に列記されている遺伝子から、SNV-1(韓国人サンプル中の少なくとも1名が有する韓国人固有の遺伝子変異)が数量的に多い遺伝子について、グーグル論文検索で関連する疾患を個別検索すると、肉体面よりも明らかに何らかの精神疾患との関連性を示す遺伝子が極めて多い
②The National Center for Biotechnology Information=アメリカ国立生物工学情報センター にて、例えば、統合失調症と検索し、統合失調症関連遺伝子としてNCBIが表示してくる遺伝子名とadditional_file_7 との突合結果は下記一覧のとおり。
| 境界性障害 | 双極性障害 | シナプス結合 | 知性 | ニューロン | シナプス | 統合失調症 | うつ病 | 自閉症 | 肝臓 対比用 |
|
| gene | TPH2 | APOE | CADM1 | INTLQ2 | APOE | APOE | DAOA | CEDORA | APOE | HNF1A |
| SNV1 | 89 | 10 | 121 | 0 | 10 | 10 | 8 | 0 | 10 | 13 |
| gene | GRIN1 | CACNA1C | CDH1 | INTLQ3 | PRKN | DLG1 | CACNA1C | GRM7 | AUTS2 | ALPL |
| SNV1 | 18 | 302 | 44 | 0 | 0 | 91 | 302 | 379 | 420 | 31 |
| gene | SLC6A4 | ACE | ICAM1 | INTLQ1 | APP | DLG4 | TCF4 | ATP6V1B2 | ACE | SLCO1B1 |
| SNV1 | 18 | 18 | 10 | 0 | 124 | 16 | 147 | 17 | 18 | 46 |
| gene | CDH2 | DAOA | CDH2 | NBN | SMN1 | NRXN1 | MIR137 | SLC18A1 | UGT1A1 | APOE |
| SNV1 | 78 | 8 | 78 | 28 | 6 | 418 | 27 | 29 | 68 | 10 |
| gene | COMT | HLA-E | PTK2B | RXRA | REST | BDNF | HLA-E | SP4 | HLA-B | HLA-B |
| SNV1 | 8 | 0 | 57 | 79 | 14 | 30 | 0 | 38 | 3 | 6 |
| gene | ACE | MAD1L1 | ACE | COL1A2 | NRG1 | DLG2 | Disc1 | LZTS1 | BDNF | ABO |
| SNV1 | 18 | 287 | 18 | 16 | 388 | 789 | 144 | 27 | 30 | 26 |
| gene | MTOR | WWOX | ITGB1 | PAX5 | SMN2 | SHANK3 | RGS4 | MKLN1 | PTEN | ACE |
| SNV1 | 41 | 797 | 33 | 73 | 5 | 35 | 7 | 104 | 30 | 18 |
| gene | FKBP5 | BDNF | NRXN1 | LRPPRC | BDNF | PRKN | ANK3 | LY86 | PRKN | PNPLA3 |
| SNV1 | 45 | 30 | 418 | 56 | 30 | 0 | 229 | 36 | 0 | 20 |
| gene | DRD4 | KIT | L1CAM | PRMT6 | PCSK9 | CTNNB1 | AS3MT | LY86-AS1 | PCSK9 | GLS2 |
| SNV1 | 21 | 24 | 4 | 4 | 12 | 19 | 17 | 123 | 12 | 7 |
| gene | GRIN2B | GNA12 | CADM2 | DEPDC5 | UCHL1 | COMT | MAD1L1 | LINC00578 | COMT | AGT |
| SNV1 | 137 | 54 | 471 | 47 | 8 | 8 | 287 | 115 | 8 | 9 |
| gene | KIT | COMT | CADM3 | GYPC | SEMA3A | DRD2 | CSMD1 | ITGB1 | ESR1 | UGT1A1 |
| SNV1 | 24 | 8 | 17 | 26 | 216 | 20 | 1786 | 33 | 146 | 68 |
| gene | PDGFRB | ANK3 | APP | TUSC3 | ENO2 | APP | DISC2 | HOMER1 | HLA-A | KDR |
| SNV1 | 16 | 229 | 124 | 220 | 1 | 124 | 0 | 52 | 13 | 19 |
| gene | BAP1 | APOB | NCAM1 | CNTN4 | CTNNB1 | MAPT | NT5C2 | RORA | MECP2 | ALDH2 |
| SNV1 | 6 | 20 | 102 | 485 | 19 | 33 | 26 | 257 | 11 | 18 |
| gene | PRKCA | FADS1 | SNCA | AOX1 | ASCL1 | SHANK2 | DPYD | DMD | HFE | CETP |
| SNV1 | 195 | 15 | 42 | 33 | 7 | 322 | 319 | 373 | 18 | 17 |
| gene | CLU | FUT2 | DLG4 | ZNF365 | GDF5 | DLG3 | SCZD1 | ADM | HERC2 | STK11 |
| SNV1 | 9 | 13 | 16 | 102 | 16 | 10 | 0 | 471 | 131 | 19 |
| gene | CSMD1 | SYNE1 | DRD2 | UCN3 | PTEN | CXCR4 | SCZD2 | ANXA1 | RYR2 | NAT2 |
| SNV1 | 1786 | 170 | 20 | 78 | 30 | 0 | 0 | 12 | 304 | 3 |
| gene | DPYD | ESR1 | NECTIN1 | PISD | TP53 | WNT5A | ITIH3 | VCAN | NRXN1 | CPT1A |
| SNV1 | 319 | 146 | 0 | 21 | 12 | 9 | 13 | 36 | 418 | 41 |
| gene | HDAC4 | FCGR2A | SYP | ESF1 | PSEN1 | ATXN3 | SCZD6 | CDH13 | MTHFR | SLCO1B3 |
| SNV1 | 191 | 24 | 2 | 21 | 28 | 20 | 0 | 600 | 13 | 147 |
| gene | NR4A2 | CSMD1 | TNF | KIF16B | BMP2 | APOA4 | SCZD10 | DDX21 | CNTNAP2 | HAMP |
| SNV1 | 9 | 1786 | 0 | 118 | 5 | 2 | 0 | 11 | 940 | 4 |
| gene | FBXO11 | TCF4 | CTNNB1 | ELSPBP1 | BCL2 | SCZD3 | SLC6A15 | PON1 | ||
| SNV1 | 55 | 147 | 19 | 16 | 73 | 0 | 26 | 118 | ||
| 合計 | 3083 | 4088 | 1596 | 1423 | 1004 | 1956 | 3312 | 2739 | 2711 | 522 |
| 1297 | 2302 | 1596 | 1423 | 1004 | 1956 | 1526 | 2739 | 2711 | 522 |
以上からして、韓国人固有の遺伝子変異は明らかに肉体面ではなく、ヒトの精神面に集中して生じてしまっている。記事筆者の推定原因は、上記大文字箇所のとおり。
(注) 2 遺伝子頻度の変動
この記述の分析が正しいと仮定した場合でも、遺伝子頻度は、人口の大幅な減少・ランダムに交配していない等の何らかの特殊事由がない限り、30世代程度(高麗時代~現在)では大きくは変動しないはずである。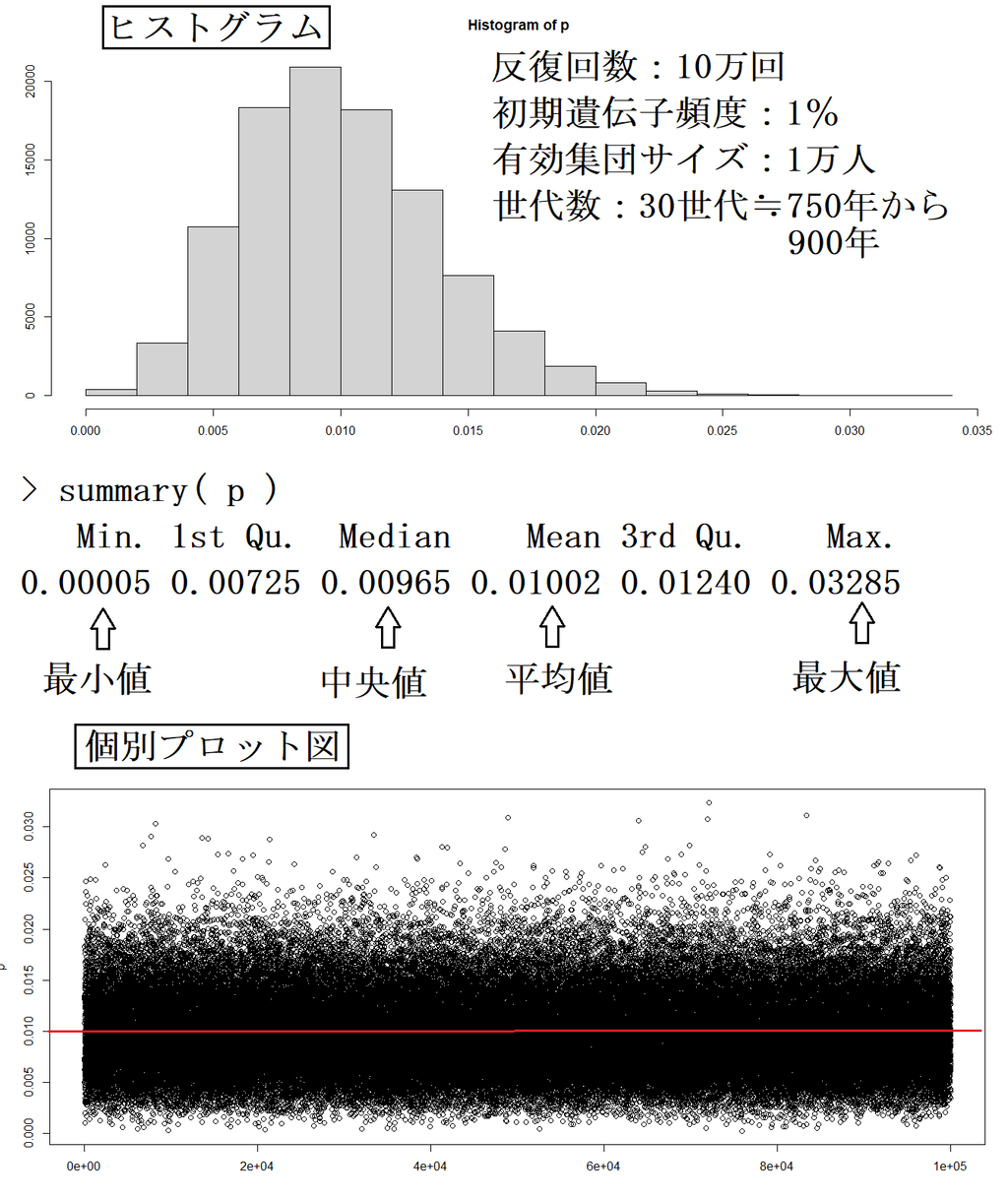
使用ソフト=R
主たるコード内容は下記。
install.packages( "datarium" )
# data.frame to be filled
wf_df <- data.frame()
# effective population sizes
sizes <- c(10000)
# starting allele frequencies 1%
starting_p <- c(0.01)
# number of generations
n_gen <- 30
# number of replicates per simulation
n_reps <- 100000
# run the simulations
for(N in sizes){
for(p in starting_p){
p0 <- p
for(j in 1:n_gen){
X <- rbinom(n_reps, 2*N, p)
p <- X / (2*N)
rows <- data.frame(replicate = 1:n_reps, N = rep(N, n_reps),
gen = rep(j, n_reps), p0 = rep(p0, n_reps),
p = p)
}
}
}
wf_df <- rbind(wf_df, rows)
plot( p )
【参考】有効集団サイズが小さい場合には、遺伝子頻度1%の変異は、消滅する可能性が高い(集団遺伝学の理論上の値である選択係数は、考慮していない)

(注) 3 集団遺伝学論文による近親相姦の子に出てしまう影響
論文 6
Extreme inbreeding in a European ancestry sample from the contemporary UK population
Loic Yengo et.al
Nature communications 03 September 2019
①近親相姦の影響について、バイオバンクUKの約35万人のDNAデータ(分析対象SNP数は約30万に過ぎない)から、125人の近親相姦例を検出した上記論文によれば近親相姦の影響が出ている形質は、次のとおり。P値による判定は、0.05ではなく、0.005でなされており、次の形質(P値順)が近親相姦と関連するとしている
*第一位:肺機能(肺活量ではなく、最大呼気流速度)
*第二位:身長
*第三位:握力
*第四位:流動性知能(内容は、下の表)
*第五位:同じ内容に対する反応時間測定(具体的な内容は、下の表 )
*第六位:聴力
*第七位:学歴(教育を受けた年数)
論文 6 表3より
下の表はこの論文より
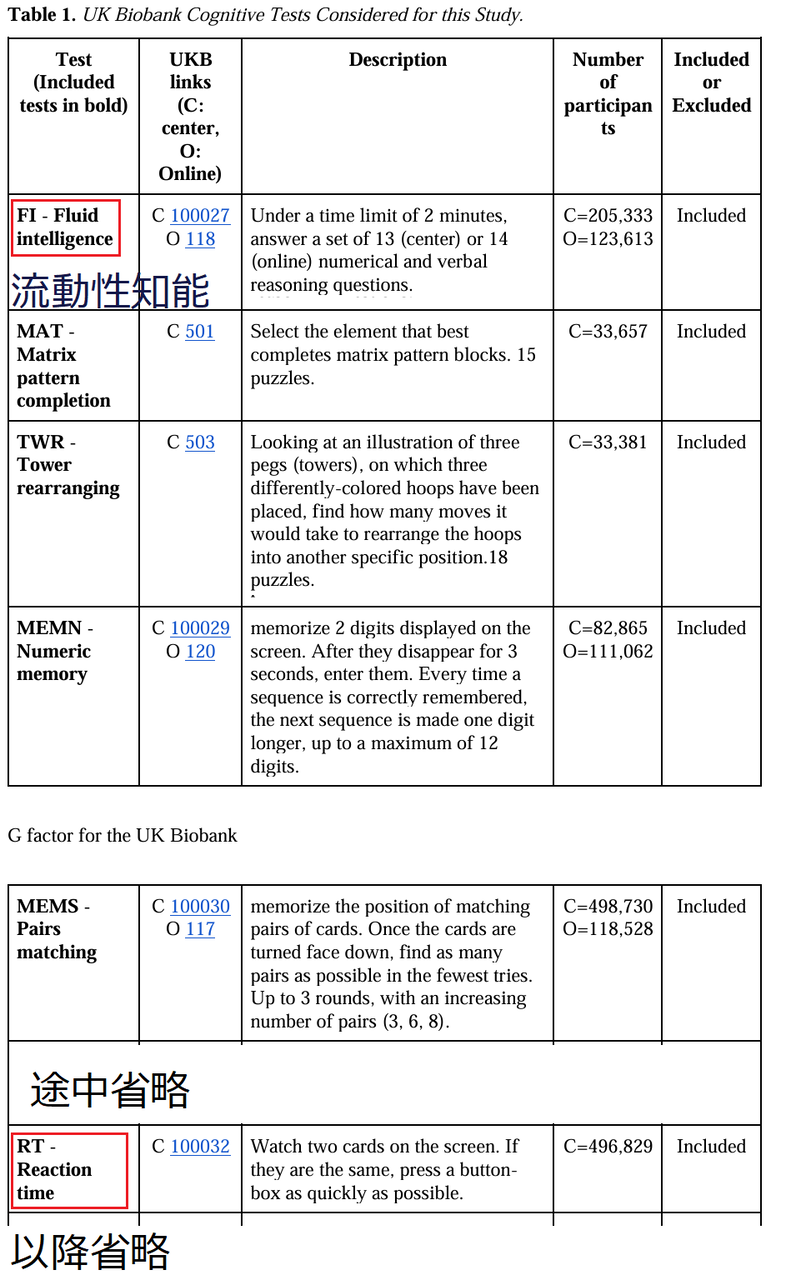
②バイオバンクUKデータには、ゲノムデータだけではなく、調査協力者の身長・非常に簡単なIQテスト類似の認知能力関連測定結果・子供の数等々のデータが含まれている。ただし、バイオバンクUKの形質データは、全イギリス人平均よりも若干上方にずれていることを指摘した論文が存在する。
③上記論文 6 は、近親相姦を本人申告、又は、性的虐待による警察等関与によるデータではなく、SNP分析から個体ごとのFROHの値(注4)が0.1以上をもって、極度の近親交配=近親相姦と見做したものであり、0.03%という近親相姦比率を算出している。
論文 7
Children of incest
Patricia A. Baird et al.
November 1982
The Journal of P E D I A T R I C S
当事者申告、又は、警察等関与による実際の近親相姦例については、医師(MD=医学博士=日本と異なる)による上記の古い論文で様々な現実の疾患例が紹介されています。
近親相姦例29の結果を一言で言えば、「何が起きるかわからない」 という点に尽きるものの、29例中9例は正常です。
全ての症例に関しては、上記論文抜粋メモ
(注) 4 ROH線分図
ROH線分図には、①FROH線分図(比率)②SROH線分図(ROHの長さに応じて数値合計)③NROH線分図(一定以上の長さのROHの数)の3タイプがあり、本文掲載図は①のタイプのいわゆるバイオリンプロット図。下記がROHそのもののイメージ図。
なお、本文に掲載のFROH線分図に関しては、何らかの画像改変又はデータ操作がなされた可能性が極めて高いと記事筆者は考えている。(=赤矢印の箇所は、現実にはもっと上方ではなかろうか?と見做している。)
(注)5 両班階層が宿していた遺伝的特異性の朝鮮人集団全体への拡散
本文中の、「意図せざる同父異母の者同士の近親交配=近親相姦」は、当然ですが、SNP(=DNA)中の非同義置換変異比率を上昇させることは通常ありません。 単に、ホモ接合型比率(=ROH)を上昇させるが故に、ヘテロ接合であれば具現しない形質(例えば知能低下)や疾患(例えばアミノ酸代謝異常)が、表現型として現れるに過ぎません。 血液型=表現型がAであっても、AA(ホモ接合)の場合とAO(ヘテロ接合)の者がいるのと基本的には同じです。
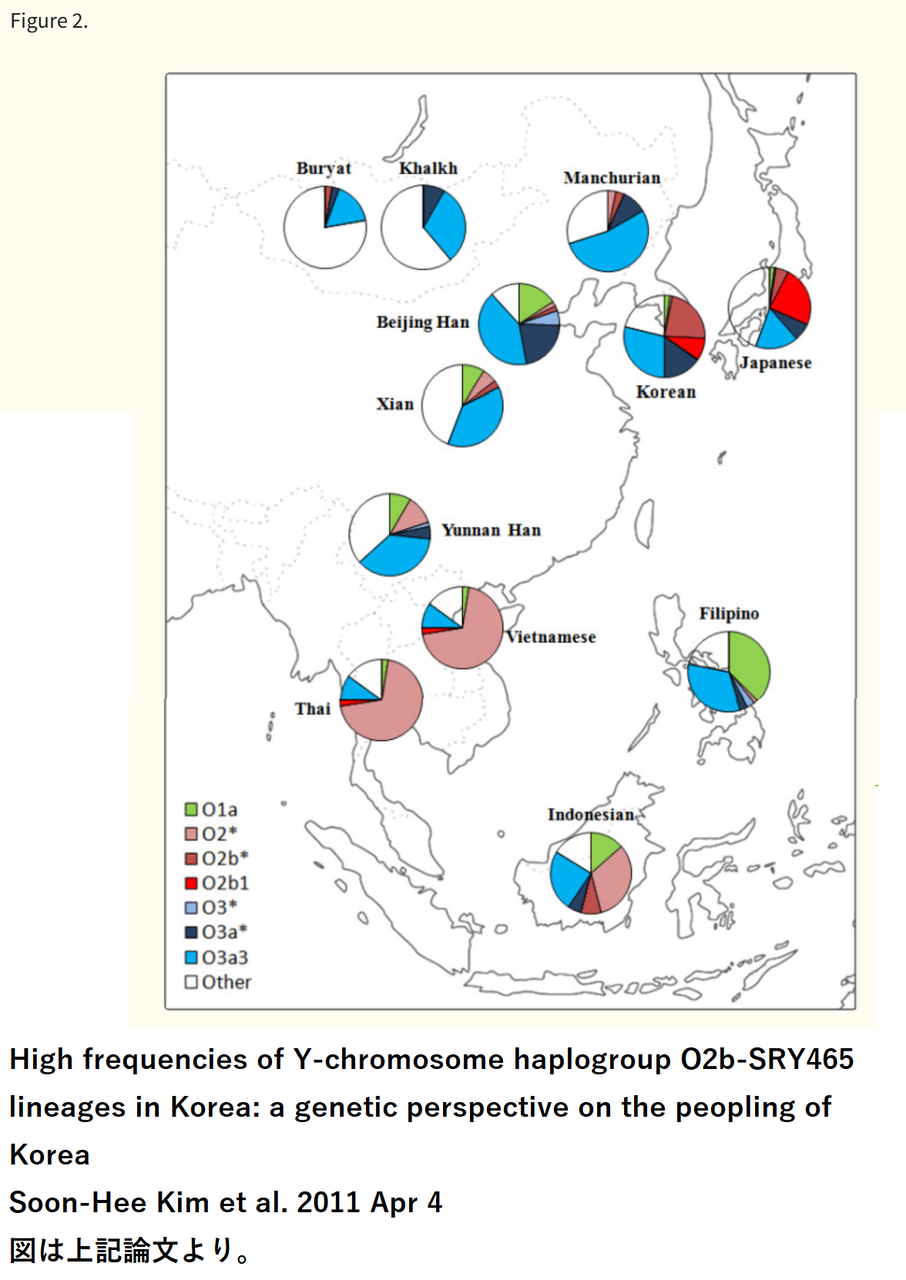
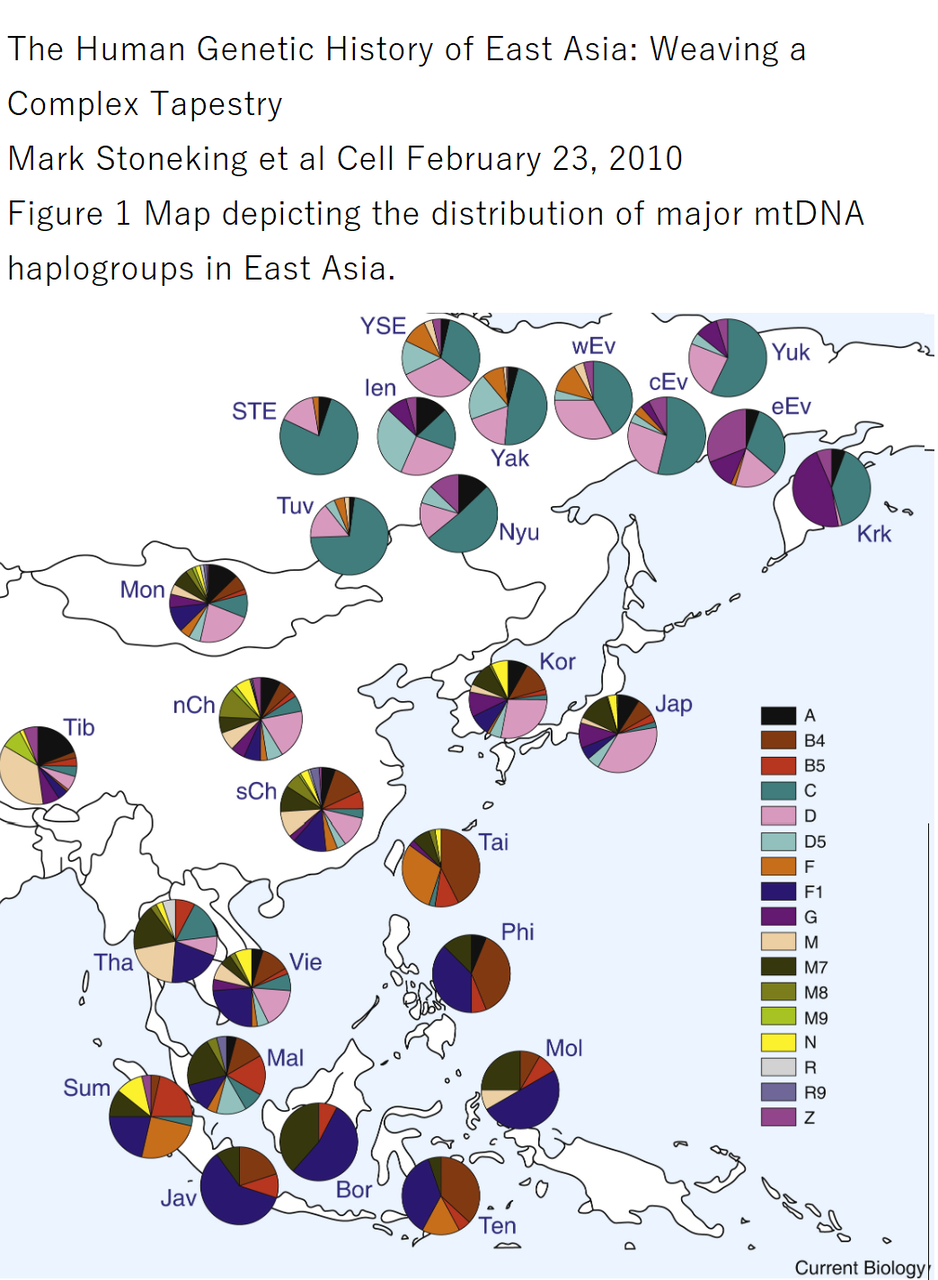
このため、13世紀頃以降の朝鮮半島で、ヒトのメンタルに関連する遺伝子の高い非同義変異比率を有していた特定の集団即ち両班階層の遺伝的特異性が 朝鮮人集団全体に広がったのであろうと推測しています。そのことを明確に示すデータは存在しないのですが、父系祖先を辿り得るY染色体ハプログループ に関し、朝鮮人男性はO2b(現在の区分でO1b2)に高い集積を示すという特質を有しています。(=下の図のO2bの比率が高麗末期以降約700年の間に、朝鮮独特の同胞奴隷制により徐々に高まったという趣旨です) 母系祖先を辿り得るミトコンドリアDNAに関しては、朝鮮人の場合、特徴的な分布を示してはいません
Y染色体ハプログループ
ミトコンドリアDNA
この投稿の元内容及び他の投稿に興味がある方は、下記サイトをご覧ください
アメーバブログのみの追記(遺言)
論文⑤A global reference for human genetic variation The 1000 Genomes Project Consortium
The 1000 Genomes Project Consortium
Nature 2015 Sep 30 より
